 Setting up apparatus
Setting up apparatus Video capture & tracking
Video capture & tracking Observing behaviour
Observing behaviour Connecting equipment
Connecting equipment Automating complex tests
Automating complex tests Running tests
Running tests Results
Results Visualising data
Visualising data Analysis
Analysis Transferring data
Transferring data Open field
Open field Water-maze
Water-maze Y-maze
Y-maze Fear conditioning
Fear conditioning Novel object
Novel object Barnes maze
Barnes maze Radial arm maze
Radial arm maze Light/dark box
Light/dark box Operant conditioning
Operant conditioning Zebrafish
Zebrafish Computers
Computers Multifunction remote
Multifunction remote Accessories
Accessories Digital interface
Digital interface Optogenetic interface
Optogenetic interface Synchronisation interface
Synchronisation interface Relay interface
Relay interface Audio interface
Audio interface Touch interface
Touch interface Analogue interface
Analogue interface USB TTL cable
USB TTL cable Animal shocker
Animal shocker Components
Components Place preference
Place preference ANY-box
ANY-box T-maze
T-maze Zero maze
Zero maze Hole board
Hole board Sociability cage
Sociability cage OPAD
OPAD RAPC
RAPC Waterwheel forced swim test
Waterwheel forced swim test Thermal gradient ring
Thermal gradient ring Operon
Operon Activity Wheel
Activity Wheel Full ANY-maze licence
Full ANY-maze licence Other licence types
Other licence types Developing countries licence
Developing countries licence Contact support
Contact support Support Policy
Support Policy FAQs
FAQs Guides
Guides Downloads
Downloads Send us files
Send us files Activate a licence ID
Activate a licence ID Contact us
Contact us Blog
Blog About
About Testimonials
Testimonials Privacy Policy
Privacy Policy
Automation of the Morris water-maze test requires robust tracking, flexible zone design, good experiment management and a selection of specialist results
ANY-maze provides all these features and more. Use the tabs below to learn more about using ANY-maze with the Morris water-maze.
We've included some videos of real water-maze tests, so you can see the system in action, as well as details on recommended equipment, such as suitable cameras, and a list of results that are especially useful in this test.

Tracking
Tracking in the water-maze poses a unique set of challenges including reflections from the water, instances when the animal dives and the wide variation in conditions under which the test is performed.
The video on the right shows ANY-maze tracking an animal in a black maze in very low light. Note the reflection in the top-right, which, while not severe, is very similar in shape and colour to the rat. Despite these challenges, ANY-maze tracks without errors.

Starting tests
Another common requirement in the water-maze is for tracking to start as soon as the animal is placed in the maze. In ANY-maze this can be achieved using the auto-start option, which automatically begins the test as soon as the experimenter leaves the camera’s view – as demonstrated in this video.
In some cases this may not be fast enough – for example, if the experimenter has to walk around the apparatus in order to leave the camera’s view – in which case you can use an ANY-maze remote start switch, to begin the test the instant the animal enters the water.

Moveable zones
The water-maze usually includes an escape platform, but this is often placed in a different location for each animal. In ANY-maze it’s easy to create moveable zones together with a list of the different locations they can adopt – see the image in the right.
Once the platform zone is set up, you just have to tell ANY-maze where it will be located for each animal and the software does the rest – automatically taking the location into consideration when determining test end, calculating results, etc.

Dividing the experiment into stages
Water-maze experiments are almost always divided into different stages, for example a training stage and a probe stage. ANY-maze allows you to specify the experiment design so it is aware of the different stages and the trials within them.
This means that while running the tests ANY-maze can automatically assess such things as training criteria, and it also helps when analysing results, as you can easily perform comparisons between different groups in different stages and/or trials.
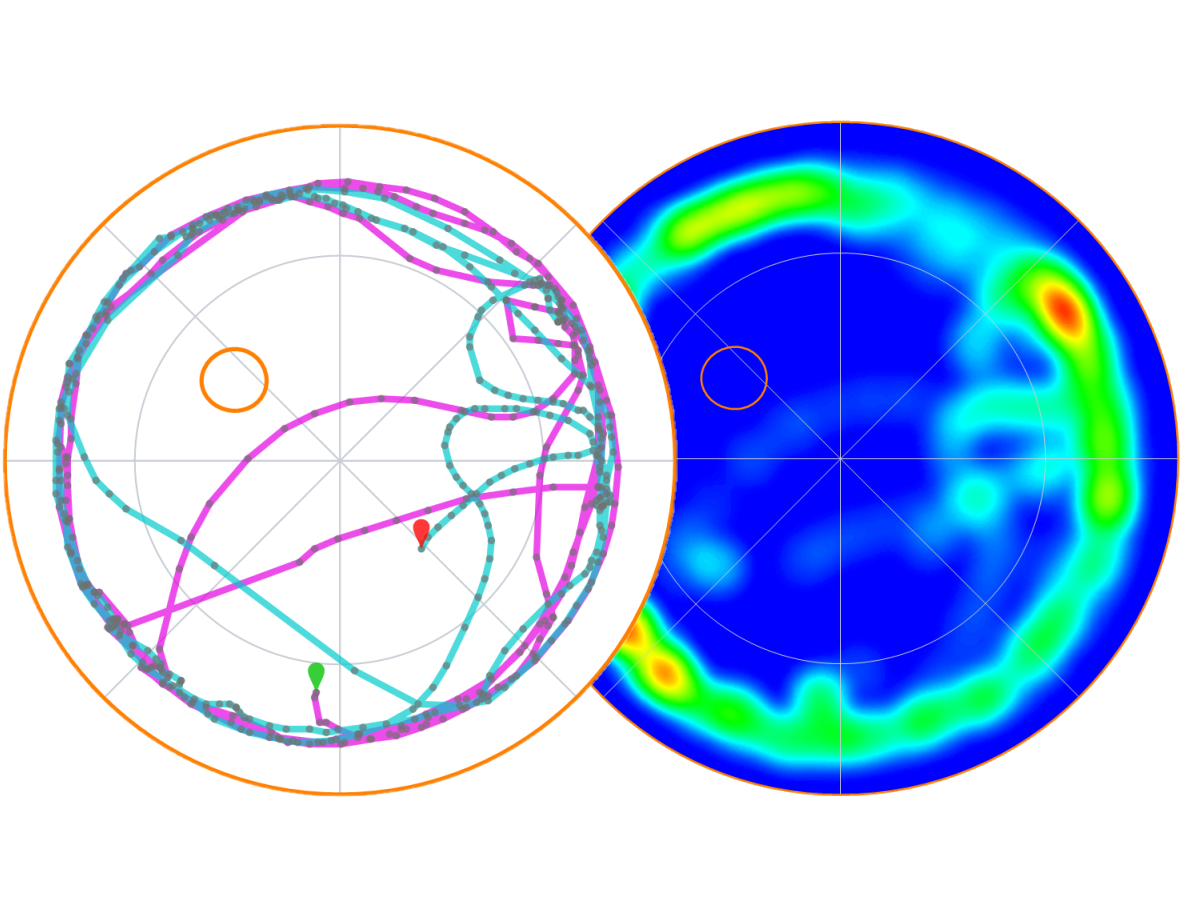
Track plots and heat maps
Track plots and heat maps provide excellent tools for visualising the animals’ search strategies in the water-maze. Track plots can be colour coded, for example, by time segment, and can include markers to indicate where specific events occurred.
Heat maps can show data for an individual test, but can also show mean data for groups of animals – which can be very useful for visually comparing the groups’ performance in probe trials.
Both formats can easily be copied and pasted into presentations or saved in high resolution for publication.
Nothing’s set in stone
In ANY-maze almost everything defined in the protocol can be altered before, during or even after an experiment has been performed.
For example, you might perform a water-maze experiment without including a Thigmotaxis zone – but this wouldn’t prevent you from adding one afterwards. ANY-maze would simply re-analyse the tracking data and almost instantly report the results for the new zone – watch the video to see this in action.

Results
ANY-maze can provide literally hundreds of results for any test, but some of those that are commonly used in the water-maze include:
- Latency to enter the platform zone
- Distance travelled before entering the platform zone
- Time in the platform zone quadrant
- Time in any defined zone, for example a thigmotaxis zone
- Time in Whishaw’s corridor
- Distance travelled in Whishaw’s corridor
- Corrected integrated path length (CIPL)

Tracking a rat in a water-maze
in low light
In this example a white rat is tracked in a dark coloured maze. The lighting was low (around 3 lux), but a sensitive camera was used so the apparatus appears to be well lit.

Tracking a white rat in a white
water-maze
Another real-world example of tracking, this time a white rat is tracked in a white water-maze, so there’s little contrast between the animal and the background.

Tracking in a black water-maze in
very low light
This example pushes things to the limit – a white rat is tracked in a dark coloured maze, but in extremely low light.

Mazes
We manufacture our own water-maze, which is available in three diameters. The maze is mounted on a wheeled cart which provides convenience in positioning and storing, as well as elevating the maze making it easier to use and to drain. The cart has swivel casters with wheel brakes.
View more
Cameras
USB camera
The ANY-maze USB camera is an excellent choice for the water-maze. We recommend fitting this camera with a varifocal (zoom) lens, so you can simply mount the camera on the ceiling and then zoom in and out until the maze nicely fits the camera's view.
View more
Webcam
A webcam provides an inexpensive alternative to the ANY-maze camera. If you intend to test in normal lighting conditions (>= 100 lux) and you can mount the camera far enough from the maze for it to see it all, then a webcam should work well. Another potential issue is the cable length; most webcams have quite short cables.

Lighting
Visible light
You should try to avoid direct lighting over the maze. A good solution is to shine lights up at the sides of the room so the light reflects off the ceiling - this usually creates uniform difuse illumination over the maze and reduces reflections.

Infrared illuminator
If you intend to track in low light (< 10 lux) or in darkness, then you will do best to use an infrared illuminator and an infrared sensitive camera (most cameras are IR sensitive). Again, pointing the illuminator at the ceiling - so the light reflects down onto the maze - usually reduces reflections.
View more
Accessories
ANY‑maze Multifunction remote
The ANY‑maze Multifunction remote provides a convenient way to start the test as soon as the animal is in the maze. This remote works through walls and can be used to control four apparatus independently. It can also be used to record ambient light, temperature (for example, the temperature of the water in the maze), and sound (including ultrasound) alongside your test data.
The ANY-maze Multifunction remote
Wired remote control
The ANY-maze wired remote control provides a convenient way to start the test as soon as the animal is in the maze. This remote connects to a USB port on the PC and is water-proof, making it ideal for use with wet hands.
View more
Achilly, N.P., Wang, W. & Zoghbi, H.Y. (2021) Presymptomatic training mitigates functional deficits in a mouse model of Rett syndrome. Nature 592, 596–600
Vijayan M, Bose C, Reddy PH. (2021) Protective effects of a small molecule inhibitor, DDQ against amyloid beta in Alzheimer’s disease. Mitochondrion. 2021 Apr 9;59:17-29. doi: 10.1016/j.mito.2021.04.005.
Xie L, Fang Q, Wei X, Zhou L, Wang S. (2021) Exogenous insulin-like growth factor 1 attenuates sevoflurane anesthesia-induced cognitive dysfunction in aged rats. J Neurophysiol. 2021 May 5. doi: 10.1152/jn.00124.2021.
Rashid H, Ahmed T. (2021) Gender dimorphic effect of dopamine D2 and muscarinic cholinergic receptors on memory retrieval. Psychopharmacology (Berl). 2021 Apr 23. doi: 10.1007/s00213-021-05847-2.
Tucker LB, Fu AH, McCabe JT. (2021) Hippocampal-Dependent Cognitive Dysfunction following Repeated Diffuse Rotational Brain Injury in Male and Female Mice. J Neurotrauma. 2021 Apr 28. doi: 10.1089/neu.2021.0025
Ramandi D, Elahdadi Salmani M, et al. (2021) Pharmacological upregulation of GLT-1 alleviates the cognitive impairments in the animal model of temporal lobe epilepsy. PLoS One. 2021 Jan 28;16(1):e0246068. doi: 10.1371/journal.pone.0246068.
Stazi M, Wirths O. (2021) Chronic Memantine Treatment Ameliorates Behavioral Deficits, Neuron Loss, and Impaired Neurogenesis in a Model of Alzheimer’s Disease. Mol Neurobiol. 2021 Jan;58(1):204-216. doi: 10.1007/s12035-020-02120-z.
Hascup KN, Findley CA, et al. (2021) Riluzole attenuates glutamatergic tone and cognitive decline in AβPP/PS1 mice. J Neurochem. 2021 Feb;156(4):513-523. doi: 10.1111/jnc.15224.
Zborowski VA, Heck SO, et al. (2021) Memory impairment and depressive-like phenotype are accompanied by downregulation of hippocampal insulin and BDNF signaling pathways in prediabetic mice. Physiol Behav. 2021 Feb 2:113346. doi: 10.1016/j.physbeh.2021.113346
Sumien N, Huang R, et al. (2020) Effects of dietary 5-methoxyindole-2-carboxylic acid on brain functional recovery after ischemic stroke. Behav Brain Res. 2020 Jan 27;378:112278. doi:10.1016/j.bbr.2019.112278
Hascup, E.R., Broderick, S.O., Russell, et al. (2019), Diet‐induced insulin resistance elevates hippocampal glutamate as well as VGLUT1 and GFAP expression in AβPP/PS1 mice. J. Neurochem., 148: 219-237. https://doi.org/10.1111/jnc.14634
P.F. Giannopoulos and D. Praticò (2018) Overexpression of 5-Lipoxygenase Worsens the Phenotype of a Mouse Model of Tauopathy. Molecular Neurobiology 2018; 55(7):5926-5936
G. Perez-Garcia et al. (2108) Chronic post-traumatic stress disorder-related traits in a rat model of low-level blast exposure; Behavioural Brain Research 2018; 340:117-125
C. Chang et al. (2018) Resveratrol protects hippocampal neurons against cerebral ischemia-reperfusion injury via modulating JAK/ERK/STAT signaling pathway in rats; Journal of Neuroimmunology 2018; 315:9-14
Y Hou et al. (2018) NAD+ supplementation normalizes key Alzheimer’s features and DNA damage responses in a new AD mouse model with introduced DNA repair deficiency; PNAS 2018; 115(8):E1876-E1885
R. Goel et al. (2018) Angiotensin II Receptor Blockers Attenuate Lipopolysaccharide-Induced Memory Impairment by Modulation of NF-κB-Mediated BDNF/CREB Expression and Apoptosis in Spontaneously Hypertensive Rats; Molecular Neurobiology 2018; 55(2): 1725-1739
T Sen et al. (2017) Activation of PERK elicits memory impairment through inactivation of CREB and downregulation of PSD95 following Traumatic Brain Injury. J Neurosci 2017; 37(24):5900-5911
NS Jardim et al. (2017) Bisphenol A impairs the memory function and glutamatergic homeostasis in a sex-dependent manner in mice: Beneficial effects of diphenyl diselenide. Toxicol Appl Pharmacol 2017;329:75-84.
X Zhou et al. (2017) Zeaxanthin improves diabetes-induced cognitive deficit in rats through activiting PI3K/AKT signaling pathway. Brain Res Bull 2017; 132:190-198.JA Stokum et al., (2017) Glibenclamide pretreatment protects against chronic memory dysfunction and glial activation in rat cranial blast traumatic brain injury. Behav Brain Res 20-17; 333:43-53
S Nurdiana et al. (2017) Improvement of spatial learning and memory, cortical gyrification patterns and brain oxidative stress markers in diabetic rats treated with Ficusdeltoidea leaf extract and vitexin. Journal of Traditional and Complementary Medicine.
JLN Sprowles et al. (2017) Differential effects of perinatal exposure to antidepressants on learning and memory, acoustic startle, anxiety, and open-field activity in Sprague-Dawley rats. Int J Dev Neurosci 2017;61:92-111
M Van Can et al. (2017) Willughbeia cochinchinensis prevents scopolamine-induced deficits in memory, spatial learning, and object recognition in rodents. J Ethnopharmacol 2017.
Lin, Yu-Lung, et al. (2017) Cellular retinoic acid binding protein 1 modulates stem cell proliferation to affect learning and memory in male mice. Endocrinology (2017).
M Van Can et al. (2017) Willughbeia cochinchinensis prevents scopolamine-induced deficits in memory, spatial learning, and object recognition in rodents. J Ethnopharmacol 2017.
Lin, Yu-Lung, et al. (2017) Cellular retinoic acid binding protein 1 modulates stem cell proliferation to affect learning and memory in male mice. Endocrinology (2017).
Kim, Brian H., et al. (2017) Age-Dependent Effects of ALK5 Inhibition and Mechanism of Neuroprotection in Neonatal Hypoxic-Ischemic Brain Injury. Developmental Neuroscience 39.1-4 (2017): 338-351.
H Ning, et al. (2017) Effects of haloperidol, olanzapine, ziprasidone, and PHA‐543613 on spatial learning and memory in the Morris water maze test in naïve and MK‐801‐treated mice. Brain Behav. 2017;e00764
Pritchett, D., Taylor, A. M., Barkus, et al. (2016) Searching for cognitive enhancement in the Morris water maze: better and worse performance in D-amino acid oxidase knockout (Dao-/-) mice. European Journal of Neuroscience, 43: 979–989. doi: 10.1111/ejn.13192
Tomer Illouz, Ravit Madar, Yoram Louzoun, et al. (2016) Corrigendum to “Unraveling cognitive traits using the Morris water maze unbiased strategy classification (MUST-C) algorithm” [Brain Behav. Immun. 52C (2016) 132–144]
Au J. L., Weishaupt, N., Nell, H. J., et al. (2016) Motor and Hippocampal Dependent Spatial Learning and Reference Memory Assessment in a Transgenic Rat Model of Alzheimer’s Disease with Stroke. J. Vis. Exp. (109), e53089, doi:10.3791/53089 (2016).
Giuseppe D’Agostino, Claudia Cristiano, David J. Lyons, et al. (2015) Peroxisome proliferator-activated receptor alpha plays a crucial role in behavioral repetition and cognitive flexibility in mice, Molecular Metabolism, Volume 4, Issue 7, July 2015, Pages 528-536, ISSN 2212-877