 Setting up apparatus
Setting up apparatus Video capture & tracking
Video capture & tracking Observing behaviour
Observing behaviour Connecting equipment
Connecting equipment Automating complex tests
Automating complex tests Running tests
Running tests Results
Results Visualising data
Visualising data Analysis
Analysis Transferring data
Transferring data Open field
Open field Water-maze
Water-maze Y-maze
Y-maze Fear conditioning
Fear conditioning Novel object
Novel object Barnes maze
Barnes maze Radial arm maze
Radial arm maze Light/dark box
Light/dark box Operant conditioning
Operant conditioning Zebrafish
Zebrafish Computers
Computers Multifunction remote
Multifunction remote Accessories
Accessories Digital interface
Digital interface Optogenetic interface
Optogenetic interface Synchronisation interface
Synchronisation interface Relay interface
Relay interface Audio interface
Audio interface Touch interface
Touch interface Analogue interface
Analogue interface USB TTL cable
USB TTL cable Animal shocker
Animal shocker Components
Components Place preference
Place preference ANY-box
ANY-box T-maze
T-maze Zero maze
Zero maze Hole board
Hole board Sociability cage
Sociability cage OPAD
OPAD RAPC
RAPC Waterwheel forced swim test
Waterwheel forced swim test Thermal gradient ring
Thermal gradient ring Operon
Operon Activity Wheel
Activity Wheel Full ANY-maze licence
Full ANY-maze licence Other licence types
Other licence types Developing countries licence
Developing countries licence Contact support
Contact support Support Policy
Support Policy FAQs
FAQs Guides
Guides Downloads
Downloads Send us files
Send us files Activate a licence ID
Activate a licence ID Documentation
Documentation Contact us
Contact us Blog
Blog About
About Testimonials
Testimonials Privacy Policy
Privacy Policy
The open field test is about emotionality and locomotion, so automating it requires adaptable zone design and a comprehensive selection of locomotor related results.
Automating the open field test with ANY-maze is straightforward and provides results that would be difficult, if not impossible, to determine with simple observation and hand scoring - see the Benefits tab, below, to learn more.
On the other tabs you'll find videos of open field tests, recommended equipment and a list of results that are especially useful in this test.

User defined zones
In ANY-maze you define zones that are specific to how you want to perform tests in your apparatus – nothing’s prescribed.
In this example video, zones have been set up to to score how much time the animal spends in the corners, how much time close to the walls and how much time in the centre of the enclosure.
In fact, for every zone ANY-maze reports more than 60 different measures, so you’ll learn much more than just how long the animal spent in each location.


Going beyond zones
As well as scoring the animal’s behaviour in zones, ANY-maze can also detect things such as:
- Periods when the animal is immobile – i.e. not moving from place to place
- Sequences of movements between the different parts of the open field – for example, transitioning across the centre, or walking right round the border.
- What direction the animal is facing – for example, facing towards the centre or the towards the walls

Scoring behaviours software can't detect
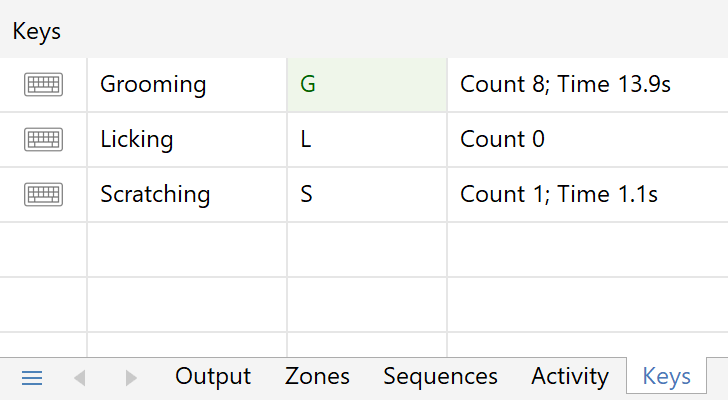
You may be interested in how much time the animals spend grooming in the open field; however this is a behaviour which software is unable to reliably detect.
To address this you can use keys to score these behaviours; for example, press the key while the animal’s grooming and release it when it stops. ANY-maze will report information such as time spent grooming and the number of grooming bouts, and it will report this separately for each zone.
Detecting rears
Rearing is another behaviour often scored in the open field but which (we believe) software is unable to reliably detect from top-down video. However, rearing can be detected using an array of photo-beams positioned such that when the animal rears it breaks beams in the array.
We manufacture an array of this type designed specifically for use with ANY-maze – it’s pictured on the right.
When this array is utilised in the open field you’ll get reports for (amongst other things) the number of rears in the different zones you define.

Viewing the animal's track
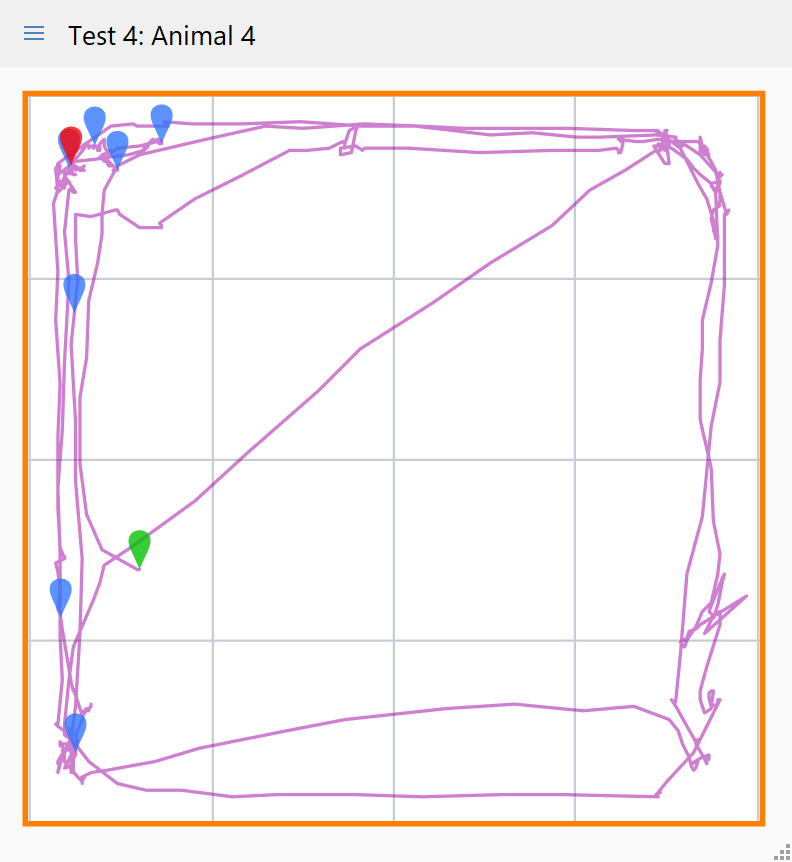
ANY-maze can plot the animal’s track or as a heat map.
Track plots, like the one on the right, show where the animal went during the test. In this case blue markers have also been included to show where the animal groomed. Perhaps unsurprisingly, it’s evident that the animal spent most of the test near the walls and mainly groomed when in the corners.
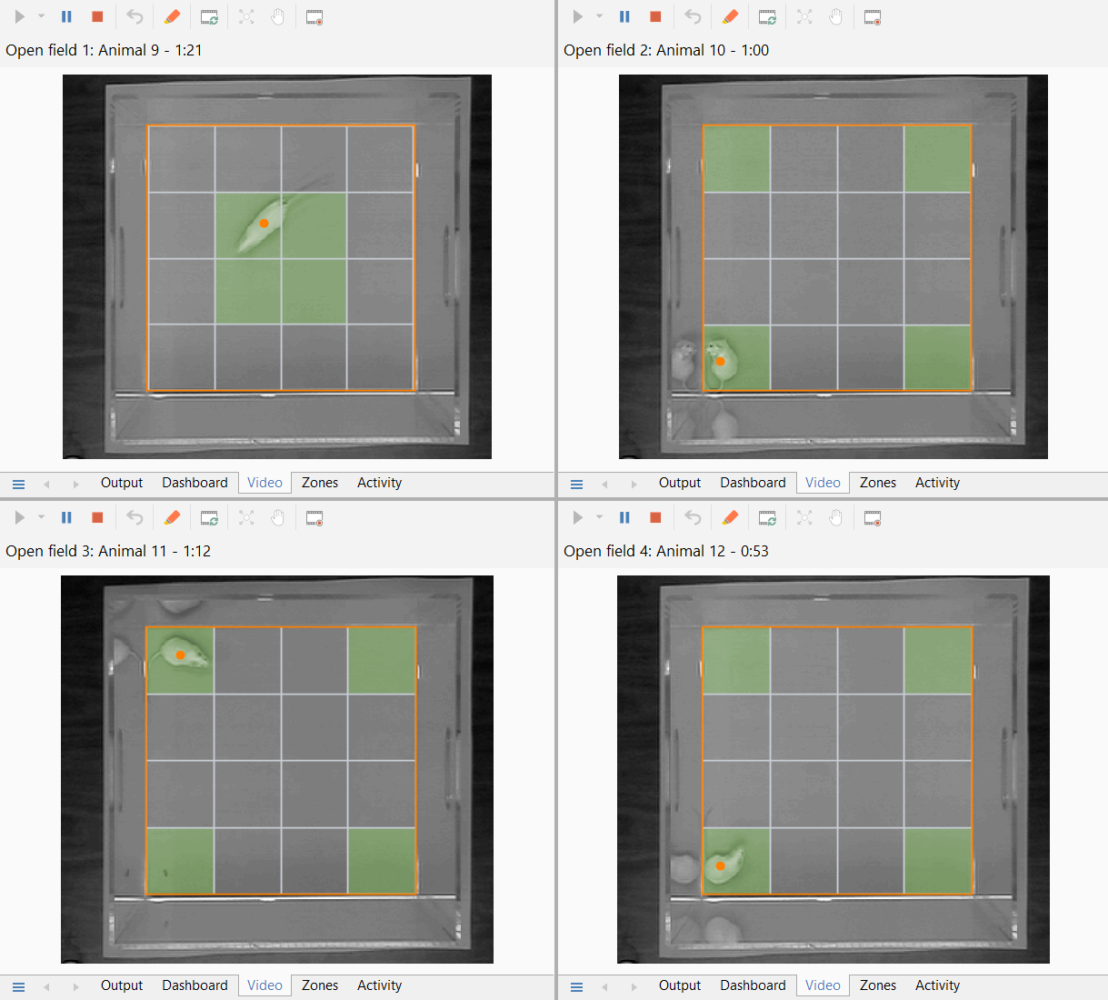
Tracking simultaneously in multiple apparatus
Tracking simultaneously in, for example, four open fields is a great way to speed up the throughput in an experiment.
- Setting up multiple open fields is hardly any more work than setting up one, as most settings are automatically applied across all the apparatus.
- You can have a single camera for each open field, or you can have multiple open fields viewed by the same camera, or any combination – for example, two cameras each viewing two open fields.
- Tests in all the apparatus can be run independently, or you can control them together if you prefer.
Almost everything can be changed
In ANY-maze you can alter the definition of a zone, a sequence or indeed almost anything else, at any time, whether before, during or after you’ve run your tests.
For example, the size of the centre zone in the open field is somewhat debatable. Some researchers minimise it, while others define it as half of the overall test arena. If you ran an experiment with a small centre zone, but then wanted to see what would change with a larger one, you could simply alter the size, and ANY-maze would immediately re-calculate the results to take this change into account.
Results
ANY-maze can provide literally hundreds of results for any test, but some of those that are particularly useful in the open field include:
- Time in the centre
- Time in the wall (thigmotaxis) zone
- Time in the corners
- Number of entries into the centre
- Distance travelled in the centre
- Distance travelled in the wall zone
- Total distance travelled
- Percentage of time in the centre
- Number of centre crossings
Traditionally the open field was often manually scored by drawing a grid onto the floor and counting the number of times the animal crossed a line. In ANY-maze you can easily add a grid to your open field and the system will automatically report line crossings, making it easy to compare data from old experiments.

Tracking in an open field in low light
In this example a white mouse was tested in an open field in very low light.
Despite the poor illumination, ANY-maze correctly tracks the animal’s head, which in this example is used to determine when the animal enters a zone.

Tracking a dark mouse in an open field with interference
In this example a dark coloured mouse was tested in an open field. Note the flashing illumination on the left side of the apparatus – despite this, ANY-maze tracks the animal correctly.

Open fields
We manufacture our own open fields, which are available for rat or mouse (the mouse version is pictured on the right). Transparent or opaque walls fit solidly into a slotted, grey, non reflective base-plate, but lift off for easy cleaning. A circular version is also available.
View more
Cameras
USB Camera
The ANY-maze USB camera is an excellent choice for the open field. We recommend fitting this camera with a varifocal (zoom) lens, so you can simply mount the camera on the ceiling and then zoom in and out until the open field nicely fits the camera's view.
View more
Web cam
A webcam provides an inexpensive alternative to the ANY-maze camera. If you intend to test in normal lighting conditions (>= 100 lux) and you can mount the camera far enough from the open field for it to see it all, then a webcam should work well. Another potential issue is the cable length; most webcams have quite short cables.

Lighting
Infrared illuminator
If you intend to track in low light (< 10 lux) or in darkness, then you may wish to use an infrared illuminator and an infrared sensitive camera (most cameras are IR sensitive).
View more
Accessories
Photobeam arrays
The ANY-maze photobeam array provides a ‘curtain’ of photobeams which can be used to detect when an animal rears. The array plugs directly into the ANY‑maze Digital interface.
The array is available both 'standalone' and as part of the ANY-box system, which includes mounting columns, and a camera.

ANY‑maze Radio remote control
The ANY‑maze Radio remote control provides a convenient way to start the test as soon as the animal is in the maze. This remote works through walls and has 2 buttons, allowing you to control two apparatus independently.
View more
Berroug, L., et al (2023). Sex-specific neurobehavioral and biochemical effects of developmental exposure to Malathion in offspring mice. Naunyn-Schmiedeberg’s Archives of Pharmacology, 42.
Krause, W.C., et al (2021). Oestrogen engages brain MC4R signalling to drive physical activity in female mice. Nature 599, 131-135
Achilly, N.P., Wang, W. & Zoghbi, H.Y. (2021) Presymptomatic training mitigates functional deficits in a mouse model of Rett syndrome. Nature 592, 596–600
Guo, L. et al (2021). Stimulation of astrocytic sigma-1 receptor is sufficient to ameliorate inflammation-induced depression. Behavioural Brain Research, 410. 113344.
Diniz, C. W. P. (2021). Classical Music and Environmental Enrichment Enhanced Spatial Memory and Learning and Increased Mouse Innate Tendency to Avoid Open Spaces. EC Neurology, 13.
Rojas-Carvajal, M., & Brenes, J. C. (2020). Acute stress differentially affects grooming subtypes and ultrasonic vocalisations in the open-field and home-cage test in rats. Behavioural processes, 176, 104140.
He, C. et al (2020). Establishing a Special Open Field Test Appliance for Tree Shrews Evaluates their Stressed Locomotor Behavior. J Vet Sci Ani Husb, 8(1), 104.
Ueno, H. et al (2019). Anti-depressive-like effect of 2-phenylethanol inhalation in mice. Biomedicine & Pharmacotherapy, 111, 1499-1506.
Madhu, L. N. et al. (2019). Neuroinflammation in Gulf War Illness is linked with HMGB1 and complement activation, which can be discerned from brain-derived extracellular vesicles in the blood. Brain, behavior, and immunity, 81, 430-443.
Zhang, X. et al (2019). The antidepressant effects of apigenin are associated with the promotion of autophagy via the mTOR/AMPK/ULK1 pathway. Molecular medicine reports, 20(3), 2867-2874.
C. Bodden et al. (2018) Evidence-based severity assessment: Impact of repeated versus single open-field testing on welfare in C57BL/6J mice; Behavioural Brain Research 2018; 336:261-268
S.M. Keller et al. (2018) Pharmacological Manipulation of DNA Methylation in Adult Female Rats Normalizes Behavioral Consequences of Early-Life Maltreatment; Frontiers in Behavioral Neuroscience 2018; 12:126
E.N. Dunn et al. (2018) Evaluating mice lacking serum carboxylesterase as a behavioral model for nerve agent intoxication; Toxicology Mechanisms and Methods 2018; 28(8): 563-572
M. Zamarbide et al. (2018) Loss of the Intellectual Disability and Autism Gene Cc2d1a and Its Homolog Cc2d1b Differentially Affect Spatial Memory, Anxiety, and Hyperactivity; Frontiers in Genetics 2018; 9:65
N. Weishaupt et al. (2018) APP21 transgenic rats develop age-dependent cognitive impairment and microglia accumulation within white matter tracts; Journal of Neuroinflammation 2018; 15:241
Sprowles, J. L. et al (2017). Differential effects of perinatal exposure to antidepressants on learning and memory, acoustic startle, anxiety, and open-field activity in Sprague-Dawley rats. International Journal of Developmental Neuroscience, 61, 92-111.
Kuniishi, H. et al (2017). Early deprivation increases high-leaning behavior, a novel anxiety-like behavior, in the open field test in rats. Neuroscience Research, 123, 27-35.
A Maple et al. (2017) Attenuated Late-Phase Arc Transcription in the Dentate Gyrus of Mice Lacking Egr3. Neural Plasticity 2017
S Vijayakumar et al. (2017) Rescue of peripheral vestibular function in Usher syndrome mice using a splice-switching antisense oligonucleotide. Hum Mol Genet.
R Longhi et al. (2017) Effect of a trans fatty acid-enriched diet on mitochondrial, inflammatory, and oxidative stress parameters in the cortex and hippocampus of Wistar rats. Eur J Nutr.
HT Nguyen, et al. (2017) Neurofibromatosis type 1 alternative splicing is a key regulator of Ras/ERK signaling and learning behaviors in mice. Hum Mol Genet 2017
J Vidal – Anuario de Psicología (2017) Individual differences in the antibody response of inbred C57Bl/6J mice and their relation to individual differences in open-field behavior. Anuario de Psicologia 2017; 47:17-22
Karl F, Grießhammer A, Üçeyler N and Sommer C (2017) Differential Impact of miR-21 on Pain and Associated Affective and Cognitive Behavior after Spared Nerve Injury in B7-H1 ko Mouse. Front. Mol. Neurosci. 10:219.
Huang, Qian, Ying Zhou, and Lin-Yun Liu. (2017) Effect of post-weaning isolation on anxiety-and depressive-like behaviors of C57BL/6J mice. Experimental Brain Research (2017): 1-7.
Y Ye et al. (2017) Alterations in Opioid Inhibition Cause Widespread Nociception but Do Not Affect Anxiety-Like Behavior in Oral Cancer Mice Neuroscience. S0306-4522(17)30443-8
K Kawabe (2017) effects of chronic forced-swim stress on behavioral properties in rats with neonatal repeated MK-801 treatment. Pharmacol Biochem Behav 2017; 159:48-54.
NR Silva et al. (2017) Antinociceptive effects of HUF-101, a fluorinated cannabidiol derivative Prog Neuropsychopharmacol Biol Psychiatry. 2017 Jul 15;79(Pt B):369-377
Stevenson, J. R., et al. (2017)Oxytocin reduces alcohol consumption in prairie voles. Physiology & Behavior.
Mammadova, Najiba, et al. (2017) Lasting Retinal Injury in a Mouse Model of Blast-Induced Trauma. The American Journal of Pathology.
S Ganapati et al. (2017) Molecular Containers Bind Drugs of Abuse in Vitro and Reverse the Hyperlocomotive Effect of Methamphetamine in Rats. Chembiochem.
WB Solecki et al. (2017) Noradrenergic signaling in the VTA modulates cocaine craving. Addict Biol. 2017
Imeh-Nathaniel, Adebobola, et al. (2017) Effects of chronic cocaine, morphine and methamphetamine on the mobility, immobility and stereotyped behaviors in crayfish.” Behavioural Brain Research. LMA
S Ganapati et al. (2017) Molecular Containers Bind Drugs of Abuse in Vitro and Reverse the Hyperlocomotive Effect of Methamphetamine in Rats. Chembiochem.
Imeh-Nathaniel, Adebobola, et al. (2017) Effects of chronic cocaine, morphine and methamphetamine on the mobility, immobility and stereotyped behaviors in crayfish. Behavioural Brain Research.
Isaksen, Toke Jost, et al. (2016) Behavior Test Relevant to a 2/a 3 Na+/K+-ATPase Gene Modified Mouse Models. P-Type ATPases: Methods and Protocols (2016): 341-351.
Curto, Yasmina, et al. (2016) Chronic benzodiazepine treatment decreases spine density in cortical pyramidal neurons. Neuroscience letters 613 (2016): 41-46.
Das, Saroj Kumar, et al. (2016) Early mood behavioral changes following exposure to monotonous environment during isolation stress is associated with altered hippocampal synaptic plasticity in male rats. Neuroscience letters 612 (2016): 231-237.
Nikiforuk, Agnieszka, et al. (2016) The effects of a 5-HT 5A receptor antagonist in a ketamine-based rat model of cognitive dysfunction and the negative symptoms of schizophrenia. Neuropharmacology 105 (2016): 351-360.
Walker, Jennifer M., et al. (2015) Beneficial effects of dietary EGCG and voluntary exercise on behavior in an Alzheimer’s disease mouse model. Journal of Alzheimer’s Disease 44.2 (2015): 561-572.